

概述
任何的體育活動,都是骨骼肌(skeletal muscles)收縮的成果,人體共有600多條骨骼肌,約佔全身重量的40%。肌肉的力量和耐力,都直接影響到運動時的表現,編排這兩方面的鍛煉時,對骨骼肌的結構和功能要有充分的認識,也就顯得非常重要了。
骨骼肌的結構
骨骼肌(在此之後只稱作肌肉)是由數以千計,具有收縮能力的肌細胞(由於其形狀成幼長的纖維狀,所以亦稱作肌纖維)所組成,並且由結締組織(connective tissue)所覆蓋和接合在一起。每一條肌纖維(亦即每一個肌細胞)均由一層稱為肌內膜(endomysium)的結締組織所覆蓋,多條肌纖維組合一起便構成了一個肌束(muscle bundle或fasciculus),並由一層稱為肌束膜(perimysium)的結締組織所覆蓋和維繫。每條肌肉可以由不同數量的肌束所組成,再由一層稱為肌外膜(epimysium)的結締組織所覆蓋和維繫。這個在肌肉內由結締組織所形成的網絡最後聯合起來,並連接到肌肉兩端由致密結締組織(dense connective tissue)構成的肌腱,再由肌腱把肌肉間接地連接到骨骼上。
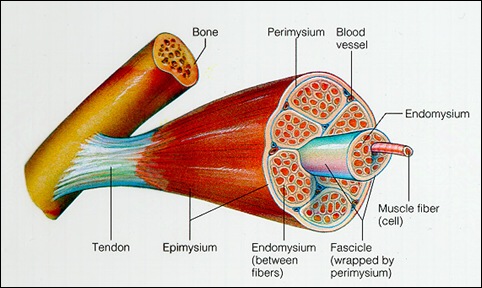
肌肉內有大量的血管和微血管,動脈和靜脈沿著結締組織進入肌肉之後,便在肌內膜之中和周圍不斷分支成更細小的血管和微血管,形成了一個非常龐大的網絡,以確保每條肌纖維都能夠得到充足的養分,與及把有害的廢物如二氧化碳等排出肌細胞之外。根據Inger(1978)及Saltin等研究員(1977),習慣坐著不動的人平均每條肌纖維只有3至4條微血管環繞著,但經常參與體育鍛煉的人卻可以有5至7條之多。
進行劇烈運動時,肌肉所需的血液可以是安靜時的100倍或以上,環繞著每條肌纖維的微血管數目當然會影響到血液的供應。除此之外,人體還會作出一些其他改變,以滿足劇烈運動時肌肉對血液供應的需求。這些改變包括:(1)活耀肌肉交替地收縮及放鬆,週期性地對血管進行擠壓,加速血液回流心臟,也就加快了血液重新供應到肌肉的速度;(2)收窄供應血液到身體非活躍部位(如內臟、腎、皮膚)的血管,另一方面卻擴張供應血液到運動肌肉的血管,以調節血液的流量。
與血管一起進入肌肉的還有神經元(即神經細胞),當中包括了運動神經元(motor neurone,亦作輸出神經元,efferent neurone)和感覺神經元(sensory neurone,亦作輸入神經元,afferent neurone)。這些神經元在結締組織內不斷分支,接觸到每條肌纖維之上。運動神經元收到來自中樞神經系統(central nervous system)的刺激後便會引起肌肉收縮。肌肉內約有60%為運動神經元,餘下來的40%為感覺神經元,主要是把痛楚和來自身體各部分的訊息傳達到中樞神經系統。
骨骼肌細胞的結構
在光學電子顯微鏡下,骨骼肌纖維(即骨骼肌細胞)呈深淺相間的橫紋,所以骨骼肌又稱作橫紋肌(striated muscle)。肌纖維膜(sarcolemma)之內是一種紅色並帶黏滯的液體,稱為肌漿(sacroplasm),當中懸浮著細胞核、線粒體、肌紅蛋白、脂肪、醣元、PC、ATP及數以千計線狀的蛋白絲,稱作肌原纖維(myofibrils)。肌原纖維內的肌節(sarcomeres),就是肌肉收縮的單位。肌節主要由兩種肌原纖維微絲(myofilaments)所組成,較幼身的一種稱作肌動蛋白微絲(actin filament),而較粗身的一種則稱作肌球蛋白微絲(myosin filament),彼此間以一種特別的結構排列著──每條肌球蛋白微絲由6條或更多的肌動蛋白微絲圍繞著。

肌原纖維的微型結構
在細心觀察之下,肌原纖維亦呈現深淺相間的橫紋,骨骼肌之所以呈深淺相間的橫紋,其實也是基於這個緣故。根據光通過肌原纖維微絲時的特性,淺色的地段稱作I帶(isotropic band),而深色的地段稱作A帶(anisotropic band)。I帶中央有一條較為深色的線,稱作Z線(zwischen line),Z線與Z線之間的一段就是一個肌節,也就是肌肉收縮的基本單位。
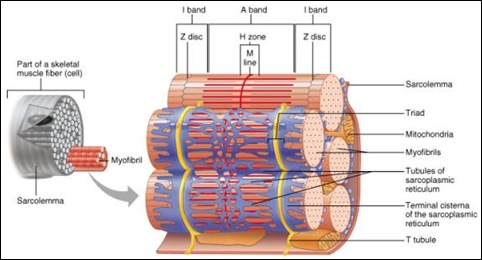
I帶只含有肌動蛋白微絲,它們在肌節之內並不延續,其中一端穩固在Z線之上,而另一端則部分伸延至A帶之內。因此,A帶雖然主要由肌球蛋白微絲所組成,但仍包含小部分的肌動蛋白微絲,A帶中央欠缺肌動蛋白微絲的部分稱作H區域(H zone)。
肌原纖維被包圍在一個稱作肌漿網(sarcoplasmic reticulum)和T小管(transverse tubules)的網絡結構之中,這個結構相信與肌肉收縮時神經訊息的傳導有關。肌漿網與T小管合共佔上肌纖維體積的5%左右,經過長期的體育鍛煉後,平均可增加至12%(Cirrito,1979)。
肌肉收縮的原理
Huxley(1969)提倡了一套微絲滑行學說(sliding filament theory),作為肌肉收縮原理的解釋。根據這套學說,肌肉收縮是由於肌動蛋白微絲在肌球蛋白微絲之上滑行所致。在整個收縮的過程之中,肌球蛋白微絲和肌動蛋白微絲本身的長度則沒有改變。
微絲滑行的實際情況仍需等待進一步的闡釋,但相信肌球蛋白微絲的突起部分(稱作橫橋或交叉橋,cross bridges)與肌動蛋白微絲上的一些特殊位置形成了一種稱作肌動肌球蛋白(actomyosin)的複合蛋白,在ATP的作用之下,就能促使肌肉產生收縮的現象。

當肌肉收縮時,若肌動蛋白微絲向內滑行,使到Z線被拖拉向肌節中央而導致肌肉縮短了,這便稱作向心收縮(亦稱作同心收縮,concentric contraction)。例如,進行引體向上(chin-up)動作時,當二頭肌(biceps)產生張力(收縮)並縮短,把身體向上提升時,就是正在進行向心收縮。反過來說,在引體向上的下降階段,肌動蛋白微絲向外滑行,使到肌節在受控制的情況下延長並回復至原來的長度時,就是正在進行離心收縮(eccentric contraction)。還有一種情況,就是肌動蛋白微絲在肌肉收縮時並未有滑動,而且仍然保留在原來位置(例如:進行引體向上時,只把身體掛在橫桿上),這便稱作等長收縮(isometric contraction)。
由於肌肉在放鬆的時候依然具有相當程度的彈性(muscle tone),所以相信此時仍有一定數量的橫橋在不斷進行工作。根據Yu與Brenner(1989),即使肌肉在放鬆的情況下,仍然可以有30%的橫橋正在執行任務。
骨骼肌的功能
骨骼肌的主要作用是進行收縮,從而產生各種不同的動作。在體育運動的領域當中,動作的質素是非常重要的,所以肌肉如何控制力量的輕重和是否所有肌纖維都具有同等功能等,都是值得關注的問題。
運動單位
人體肌纖維的數量超過二千五百萬條,但運動神經元的數量卻只有420,000條左右。因為每一條肌纖維都受到運動神經元支配,所以每一條運動神經元必須不斷分支開去,才能達至每一條運動神經纖維支配著1至多條肌纖維的比率。由於所有受同一運動神經元支配的肌纖維都會同時收縮或放鬆,亦即整體地運作,所以每一條獨立的運動神經元和所有受其支配的肌纖維被統稱為一個運動單位(motor unit),而運動單位也是骨骼肌的基本運作單位。
每一條運動神經元所支配的肌纖維數量與肌肉本身的大小並無實際關係,反而與肌肉運作時要達至的精確度和協調性有關。負責細緻和精密工作的肌肉(如眼部肌肉),每一個運動單內可能只有一條至數條的肌纖維。反過來說,專責粗重工作的肌肉(如四頭肌),每一個運動單位內就可以有數百以至數千條的肌纖維。
全或無定律
當肌肉或神經元受到刺激,而且刺激的強度夠大的時候,就會產生肌肉收縮或把神經信息傳導開去的情況;若果刺激的強度不足,肌肉或神經元便不會作出類似的反應,這個現象稱為全或無定律(all-or-none law)。由於每一個運動單位是由一條運動神經元和所有受其支配的肌纖維組成,所以運動單位亦會按照全或無定律而運作。不過,就整條肌肉而言,則不受制於全或無定律,因為在任何的一瞬間,肌肉內的運動單位,都可以部分處於收縮的狀態,而部分卻處於放鬆的狀態。
運動單位與力量漸變
力量漸變(strength gradation)的能力無論在日常生活或體育活動中都非常重要,如果缺乏了這種改變肌肉收縮力量的能力,根本就不可能產生順暢和協調的動作。例如,以相當於舉起50千克的肌肉力量來洗刷牙齒,後果將會是不敢想像。
基本上有兩種途徑可以改變肌肉收縮的力量:(1)改變在同一時間內處於收縮狀態的運動單位數目(multiple motor unit summation),和(2)改變個別運動單位收縮的頻率(wave summation)。
1. 正在收縮的運動單位數目
當來自運動神經元的刺激夠大時,個別的運動單位就會按照“全或無定律”而進行收縮。因此,肌肉收縮的力量也就可以根據同時進行收縮的運動單位數目而得以調節。此外,每個運動單位內肌肉纖維的數量和大小,也會影響到一塊肌肉實際上能夠產生的力量。
在大部分的肌肉當中,運動單位內肌纖維的數目都有差異。例如,某一塊肌肉可能包含有25個運動單位,平均每個運動單位內有200條肌纖維,但最小的一個可能只有25條肌纖維,而最大的一個卻可能有近500條肌纖維之多。假設每條肌纖維可以產生5克的力量,這塊肌肉所能產生的力量最小為1個運動單位 ´ 25條肌纖 ´ 5克 = 125克(或0.125千克),而最大一個運動單位卻能產生1個運動單位 ´ 500條肌纖維 ´ 5克=2500克(或2.5千克)的力量;當肌肉內所有的運動單位都進行收縮時,所能產生的力量便為25個運動單位 ´ 200條肌纖維 ´ 5克=25,000克(或25千克)。因此,就這塊肌肉而言,便能產生由0.125千克至25千克的力量了。
2. 運動單位收縮的頻率
一個運動單位對一個刺激(神經衝動)的反應就是產生一個抽搐(twitch),亦即在一個短暫的收縮後,伴隨著肌肉的放鬆。若這個運動單位在達至完全放鬆前又再接收到第二個刺激,前後兩個刺激所引發的牽搦便會總合起來(summate),以致這個運動單位此時所能產生的力量,要比由單一個抽搐所產生的力量為大。如果刺激重複的頻率夠密,所引發的抽搐便會總合至完全融合的程度,運動單位處於這種強直(tetanus)狀態之下,便會一直保持張力(收縮力量),直至刺激停止或疲勞出現,而強直狀態下所能產生的力量,可以是單一抽搐所產生力量的3至4倍。
不同種類的運動單位
所有骨骼肌運動單位的運作都大至相同,不過並非所有運動單位的代謝和工作能力都一樣。雖然所有運動單位,亦即所有肌纖維均可於有氧或無氧下運作,但當中一些無論在生化或生理上,都較適宜於在有氧的情況下運作,而另一些則較適宜於在無氧下工作。人體的骨骼肌纖維主要可被分為慢肌纖維(slow-twitch fibers)和快肌纖維(fast-twitch fibers)兩大類,慢肌纖維亦稱作紅肌纖維或Type I纖維;快肌纖維亦稱作白肌纖維或Type II纖維,甚至可以再被劃分為IIA,IIB和IIC三類。在每一個運動單位內的肌纖維都只會屬於同一類形,亦即是說,「快」運動單位內只包含有快肌纖維,「慢」運動單位內則只有慢肌纖維。不過,在同一塊肌肉之內,卻可以由不同數量的「快」和「慢」運動單位所組成。
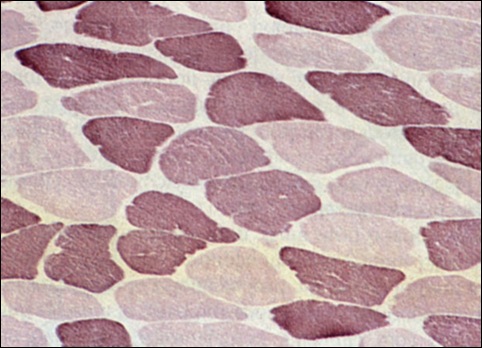
「慢」和「快」肌纖維的分佈
Vogler與Bove(1985)就曾剖驗一批死後孩童的樣本(嬰兒至8歲),並對肌纖維的分佈作出了報告和總結。早期胎兒的驅幹及四肢主要是由元始並且未曾分化的Type IIC肌纖維組成,稍後才逐漸分化成組織學上可辨認的Type I,IIA及IIB類別。各種肌纖維的成熟速率亦有所不同,例如Type I纖維在懷孕後的19至20週已經出現,但在第21至26週期間則以Type IIA及IIB肌纖維佔大多數。換句話說,在出生前,Type I肌纖維的發展是比較落後。到達36週的階段時,已經有大量的Type IIA及IIB肌纖維和小量原來的Type IIC肌纖維。因此,胎兒在子宮內的期間,所有肌纖維的成熟和分化都可說是緣於Type IIC肌纖維。
從出生至一歲,「慢」和「快」肌纖維的數目起了很大的變化。一般來說,滿一歲後,超過50%的肌纖維是屬於慢(Type I)的類形,而且「慢」和「快」肌纖維的直徑(大小)並無顯著差異。在8歲以前,男、女童肌纖維的大小亦無明顯分別。出生以後,慢肌纖維數目的激增對保持身體姿勢、運動和耐力都非常重要。當肌肉內「慢」和「快」肌纖維的比例已近均等(約各佔50%)時,生長上的主要變化只會在於肌纖維的大小方面。在正常情況下,肌纖維會於12至15歲時生長到成年人應有的大小。
在成年人的肌肉內,不同類形肌纖維的比例有很大的差異。例如,小腿的比目魚肌(soleus)就比其他腿部的肌肉多25至40%的慢肌纖維,而三頭肌(triceps)則比其他手部肌肉多10至30%的快肌纖維(Johnson,Polgar,Weightman與Appleton,1973)。不過一般來說,大部分肌肉內都混有接近相等的「慢」和「快」肌纖維。總而言之,研究結果顯示:不論(1)在同一塊肌肉的不同區域,(2)在同一個人的不同肌肉,還是(3)在不同的人體內的同一塊肌肉,肌纖維的分佈都有分別。
此外,研究結果還顯示耐力項目運動員傾向有較高百分比的慢肌纖維,而非耐力項目運動員則比沒有從事運動的人有較高百分比的快肌纖維。而且無論是否運動員,擁有較高百分比慢肌纖維的人,其最大攝氧量亦較高。其實這點亦不足為怪,因為慢肌纖維比快肌纖維的有氧代謝能力的確為高。
“訓練會否改變「慢」和「快」肌纖維的百分比”和“訓練所引致最大攝氧量的增長會否先天性地受制於「慢」肌纖維的百分比”,是兩個與肌纖維分佈有密切關係的課題。首先,大部分研究仍顯示訓練只會使到各種肌纖維無論在大小和功能上均有所增長,但顯然不會令它們彼此間互相轉換。至於第二個問題方面,肌纖維的分佈和最大攝氧量無疑是受到遺傳因素的很大限制,但研究結果亦顯示,在相同的慢肌纖維百分比之下,運動員的最大攝氧量要比非運動員為高,這也正好闡明就算是先天受到慢肌纖維百分比的限制,後天的訓練仍可顯著地提高最大攝氧量。
「慢」和「快」肌纖維在功能上的分別
慢肌纖維與快肌纖比較起來,有氧能力較高而無氧能力則較差。就算在不同類形的快肌纖維當中,Type IIA的有氧能力都要比Type IIB好,但始終仍未能高於慢肌纖維。不同類形肌纖維在結構、生化和功能上的關係見表一。
表一、不同類形肌纖維在結構與功能上的分別
|
特點 |
慢肌纖維 |
快肌纖維 |
|
|
Type I |
Type IIA |
TypeIIB |
|
|
運動神經元方面: |
|||
|
運動神經元的大小 |
小 |
大 |
大 |
|
運動神經元被激發的閾值 |
低 |
高 |
高 |
|
運動神經傳遞速度 |
慢 |
快 |
快 |
|
結構方面: |
|||
|
肌纖維直徑 |
小 |
大 |
大 |
|
肌漿網的發展 |
小 |
多 |
多 |
|
粒線體密度 |
高 |
高 |
低 |
|
微血管密度 |
高 |
中 |
低 |
|
肌紅蛋白成分 |
高 |
中 |
低 |
|
能量物質: |
|||
|
PC儲備 |
低 |
高 |
高 |
|
醣元儲備 |
低 |
高 |
高 |
|
三甘油脂儲備 |
高 |
中 |
低 |
|
酵素方面: |
|||
|
肌球蛋白-腺苷三磷酸酵素活動 |
低 |
高 |
高 |
|
醣分解酵素活動 |
低 |
高 |
高 |
|
氧化酵素活動 |
高 |
高 |
低 |
|
功能方面: |
|||
|
抽搐(收縮時間) |
慢 |
快 |
快 |
|
放鬆時間 |
慢 |
快 |
快 |
|
產生力量 |
低 |
高 |
高 |
|
能量產生的效率 |
高 |
低 |
低 |
|
抵禦疲勞的能力 |
高 |
低 |
低 |
|
彈性 |
低 |
高 |
高 |
相比之下,「慢」和「快」肌纖維的特點很多時都出現『相反』的情況。由於較細小的運動神經元有較低的刺激閾值,所以慢肌纖維會較先被徵用於活動之中。因此,慢肌纖維主要負責低強度的活動,而較高強度的運動則由Type IIA乃至是Type IIB快肌纖維負責。因為慢肌纖維包含更高密度的粒線體和肌紅蛋白,而且血液的供應亦較充足,所以比快肌纖維有較高的有氧代謝和生產ATP的能力,而且亦較耐勞。反過來說,由於快肌纖維有較高的PC和醣元儲備,而且相關的酵素活動亦較高,所以較能產生強而有力的輸出。基於種種在結構、生化和功能上的分別,快肌纖維會傾向被徵用於短時間而強度大的活動,而慢肌纖維則會被主要徵用於長時間的耐力活動。
總括來說,在一般的情況下,慢肌纖維會最先被徵用於活動之中,再視乎活動的強度、持續時間或疲勞的出現,快肌纖維亦會加入工作的行列。對於中等強度的活動,慢肌纖維和Type IIA纖維會一同運作,若活動持續下去,Type IIB纖維亦會加入工作。至於更高強度的活動,慢肌纖維和兩種快肌纖維(Type IIA和IIB)都會很快地按次序加入工作的行列。
![]()
References
![]()
Fox, E. L., Bowers, R. W, and Foss, M. L. (1993). The Physiological Basis for Exercise and Sport (5th ed.). Dubuque, IA: Wm. C. Brown.
Huxley, H. (1969). The mechanism of muscular contraction. Science, 164(3886), 1356-1366.
Inger, F. (1978). Maximal aerobic power related to the capillary supply of the quadriceps femoris muscles in man. Acta Physiol Scand, 104, 238-240.
Johnson, M. a., Polgar, J., Weightman, D., and Appleton, D. (1973). Data on distribution of fiber types in thirty-six human muscles. An autopsy study. J. Neurol Sci, 18, 111-129.
Saltin, B., Heriksson, J, Nygaard, E., and Anderson, P. (1977). Fiber types and metabolic potentials of skeletal muscles in sedentary man and endurance runners. Ann NY Acad Sci., 301, 3-29.
Vogler, C., and Bove, K. E. (1985). Morphology of skeletal muscle in children. Arch Pathol Lab Med, 109, 238-242.
![]()
Back to Anatomy & Physiology
最近更新日期(Last
Updated):2009-06-23
